
若手研究者の育成
注目の歯科基礎医学研究者!第五弾!
北海道大学大学院歯学研究院 硬組織微細構造学教室
准教授 長谷川智香
私と歯科基礎医学研究の出会いは、現所属先の主任教授であり、恩師の網塚憲生教授との出会いに始まります。将来の進路を考えていた研修医時代に網塚教授をご紹介いただき、気が付けば組織学教室所属の大学院生となって15年近くが経過しましたが、未だ気持ちは変わらず、目の前に現れた課題にひとつひとつチャレンジする日々を送っております。

研究テーマは、全身/局所リン調節因子の遺伝子改変動物を用いた骨基質石灰化/異所性石灰化メカニズムの解析や、各種骨粗鬆症治療の作用メカニズムに関する解析など、骨代謝・石灰化機序解明に関するものが中心となりますが、これらの研究は、一貫して透過型電子顕微鏡をはじめ各種顕微鏡を用いた形態学的解析を主体に進めています。形態学研究では、「組織は嘘をつかない」ため、実際の生体の中で起こっていることが視覚化される一方、研究者の力量によって本来あるはずがないもの(アーチファクト)が生まれてしまったり、視えるべきものが視えなかったりもします。組織所見から得られる真実(生体構造)を的確に捉え表現するため、当初はとにかく動物の固定から光学顕微鏡・電子顕微鏡試料の作成、切片の薄切や顕微鏡観察を繰り返し、美しい組織像を得ること、また、そこから如何にして所見を読み解いてゆくか、を繰り返しました。これら形態学解析技術・知識は、私の研究者としての基盤であり、研究を続けるうえで大きな支えになっています。一方で、近年の研究手法の発展は目覚ましく、形態学領域でも様々な顕微解析機器が開発され生物応用されているほか、様々な分子生物学的手法も進歩しています。形態学を主軸にしつつ、これら研究手法を取り入れることで、より発展的な研究にもチャレンジすることを心掛けています。「science」の世界であると同時に、「生体構造の美しさを感じる感性」の世界でもある形態学の奥深さに触れ、各々の研究を楽しみながら続けてきた結果が、現在の私を作っていることと思います。これからも、飽くことなく歯学基礎研究の世界と向き合って行ければと思います。

最後になりますが、私を研究の世界へと私を導いてくださった網塚教授、そして、共に研究を進めてきた教室の先生方、他施設の共同研究者の先生方皆さまにこの場をお借りしてお礼申し上げたいと思います。また、歯科基礎医学会は、私が初めて学会発表をさせていただいた印象深い学会であると同時に、同じ学問分野の先生方と顔を合わせ、研究や教育についてディスカッションできる大変貴重な場でもあります。このような貴重な機会を与えてくださる本会関係者の皆さまにも深く感謝申し上げます。
大阪大学大学院歯学研究科 微生物学講座
助教 大野誠之
私の高校はみな文化祭準備のために年がら学校に通っているような学校でした。3年間それを横目に高校野球に明け暮れる毎日を過ごしたことは心底痛恨の極みであったのですが、結局大学でも野球部に入りました。仲間に恵まれ、その縁もあって歯科補綴学第一教室に入局し、臨床に明け暮れました。大学院の研究は、学部生のときの基礎配属実習以来出入りしていた細菌学教室(現微生物学講座、川端重忠教授)でお世話になりました。学位取得後、そのまま微生物学講座に移住し、臨床から研究に重心を移して今に至ります。
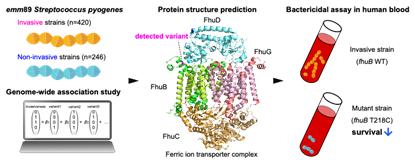
大学院では山口雅也先生にご指導いただき、ゲノムワイド関連解析による侵襲性化膿レンサ球菌感染症の発症に関与する細菌因子の探索という、バイオインフォマティクスと分子生物学的実験を組み合わせた内容を主なテーマに取り組みました。元々はin silico 解析と縁遠い場所で生きていたのですが、コロナ禍によりラボにも足を運ぶことが難しかった2ヶ月間に、家でもできる研究をと山口先生より勧められて四苦八苦しながら始めました。いまではむしろビッグデータを駆使して網羅的で面白いデータと出会うことのできるバイオインフォマティクス解析に魅せられております。
私の研究はゲノミクスが中心となっており、化膿レンサ球菌は600株以上、肺炎球菌は12,000株を超える規模で全ゲノムを収集し、致死率の高い侵襲性感染症と相関する変異や遺伝子などを網羅的に探索してきました(図)。スクリーニングののち、変異株を作製しマウス感染モデルなどを用いて病原性の証明を行いました。さらに、これらの因子を抗体医薬やワクチン抗原として応用すべく、実験を重ねています。今ではさらに研究の裾野を広げて、病原性や薬剤耐性に関連した細菌の祖先推定や進化予測についてのプロジェクトなども展開しています。兼任するバイオインフォマティクス研究ユニットでは、培った解析技術を活かして研究者や大学院生向けの講習や、臨床研究とのコラボレーションをおこなっています。
実験とin silico の解析とで座っている時間が長いため、なるべくランニング、筋トレ、あと草野球は欠かさないようにしています。奮闘中の0歳児の育児と絶妙のバランスをとりながら、なんとか頑張ろうとしている日々です。これからも元気に、微生物学および免疫学のエキスパート集う大阪大学でみなさまに育てていただける強みを活かしつつ、おもろいことを研究していきたいと思います。
九州大学大学院 歯学研究院 口腔常態制御学講座 口腔機能解析学分野、OBT研究センター、五感応用デバイス研究開発センター
講師 實松 敬介
「口腔に存在する口の形をした分子の動きを明らかにしたい」
5基本味のうち甘味やうま味を伝える味覚受容体は、クラスC Gタンパク質共役型受容体に属し、細胞外に大きなドメインを有することが知られています。その構造は、ハエトリグサ様、すなわち上顎と下顎からなる口のような形をしており、これらが開閉することで活性化状態が決まります。味分子を捕まえる(食べる)ことで、立体構造変化がおこり、味の情報伝達が始まります(図)。歯学部出身ということもあり、口腔に存在する、口の形をしたこの受容体の活性化メカニズムに非常に興味を持ち、研究を進めております。
私は、2005年に九州大学歯学部を卒業し、同歯学府歯学専攻博士課程に進学しました。ここで二ノ宮裕三名誉教授、重村憲徳教授のもと、味覚研究に着手し、一貫して食調節と味覚受容機構に関連する分子の機能解析研究を進めてきました。2018年、九州大学五感応用デバイス研究開発センター協力教員、2020年からは、歯学究院附属OBT研究センターの独立PIとしても活動しております。
主な実験手法としましては、人工味細胞を用いた受容体機能解析系の確立により、ヒトの味覚を客観的に再現し、評価しております。また近年、スーパーコンピューターを用いて、分子動力学シミュレーションを味覚受容体解析へ導入することに成功しております。分子動力学シミュレーションは、システム中の全原子に対する運動方程式を解くことで、分子の動きを予測します。多くの研究グループが、WETの専門家とDRYの専門家による共同研究により仕事を進める中、私共は両技術を駆使し、同時進行で効率的に進めることが可能です。これらの技術を用いて、甘味受容体サブユニットTAS1R3の膜貫通領域において、細胞外側でのリガンドと結合ポケットでの相互作用が、アロステリーにより受容体の細胞内側の構造変化を誘導する過程を予測することができました (Commn Biol, 2023)。
近年、味覚受容体は、口腔だけでなく全身のあらゆる臓器に発現し、生体恒常性の維持や生体防御に関与することが示唆されていますが、そのメカニズムの多くが不明です。これらの生理現象の謎に味覚受容体を切り口に迫って参りたいと考えております。
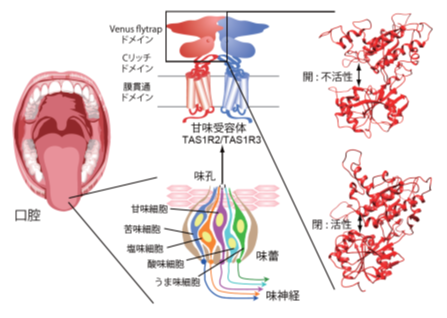
図 舌の乳頭部、口蓋、喉頭蓋、咽頭および喉頭の一部にタマネギ様の味蕾が存在する(左、中)。味蕾は、各5基本味を担当する味覚受容体を発現する各味細胞から構成される。甘味細胞はヘテロ二量体からなる甘味受容体TAS1R2/TAS1R3を発現する(中)。各サブユニットは、Venus flytrap (ハエトリグサ様) ドメイン、システイン(C)リッチドメイン、膜貫通ドメインからなる。Venus flytrapドメインは、不活性化時は開いており、甘味物質と結合すると閉じる(右)。